Imina atau imino adalah gugus fungsi atau senyawa kimia yang mengandung ikatan rangkap karbon–nitrogen. Atom nitrogen dapat melekat pada hidrogen (H) atau gugus organik (R). Jika gugus ini bukan atom hidrogen, maka senyawa tersebut kadang-kadang dapat dirujuk sebagai basa Schiff.[1] Atom karbon atom memiliki tambahan dua ikatan tunggal
Imina ( / ɪ ˈ m ː n / atau / ˈ ɪ m ɪ n / ) adalah gugus fungsi atau senyawa kimia yang mengandung ikatan rangkap karbon - nitrogen . Atom nitrogen dapat dilekatkan ke hidrogen (H) atau grup organik (R). Jika kelompok ini bukan atom hidrogen, maka senyawa tersebut kadang-kadang dapat disebut sebagai pangkalan Schiff . [1] Atom karbon memiliki dua ikatan tunggal tambahan. [2] [3] [4] Istilah "imine" diciptakan pada tahun 1883 oleh kimiawan Jerman Albert Ladenburg . [5]
Nomenklatur dan klasifikasi
Biasanya imina mengacu pada senyawa dengan konektivitas R 2 C = NR, seperti yang dibahas di bawah ini. Dalam literatur yang lebih tua, imina mengacu pada analog aza dari epoksida. Dengan demikian, etilenimin adalah spesies cincin beranggota tiga C2H4 NH. [6]
Imina terkait dengan keton dan aldehid dengan penggantian oksigen dengan kelompok NR. Ketika R = H, senyawa tersebut adalah imina primer, ketika R adalah hidrokarbil , senyawa tersebut adalah imina sekunder. Imina menunjukkan reaktivitas yang beragam dan biasanya ditemui di seluruh kimia. [4] Ketika R 3 adalah OH, imine disebut oksime , dan ketika R 3 adalah NH 2 imina disebut hidrazon .
Imina primer di mana C melekat pada hidrokarbil dan H disebut aldimine primer ; imina sekunder dengan kelompok seperti ini disebut aldimine sekunder . [7] Imina primer di mana C melekat pada dua hidrokarbil disebut ketimine primer ; imina sekunder dengan kelompok seperti ini disebut ketimin sekunder . [8]
N-Sulfinyl imines adalah kelas khusus imina yang memiliki gugus sulfinil yang terikat pada atom nitrogen.
Sintesis imina
Imina biasanya disiapkan oleh kondensasi amina primer dan aldehida dan keton kurang umum:
Lebih banyak metode khusus
Beberapa metode lain ada untuk sintesis imina.
 Reaksi yang paling penting dari imina adalah hidrolisis mereka ke senyawa amina dan karbonil yang sesuai. Jika tidak, gugus fungsi ini berpartisipasi dalam banyak reaksi lain, banyak yang analog dengan reaksi aldehida dan keton.
Reaksi yang paling penting dari imina adalah hidrolisis mereka ke senyawa amina dan karbonil yang sesuai. Jika tidak, gugus fungsi ini berpartisipasi dalam banyak reaksi lain, banyak yang analog dengan reaksi aldehida dan keton.
Agak seperti induk amina, imina sedikit bersifat basa dan dapat berefleksi secara reversibel untuk menghasilkan garam iminium. Turunan iminium sangat rentan terhadap reduksi ke amina menggunakan transfer hidrogenasi atau dengan aksi stoikiometrik natrium cyanoborohidrida . Karena imina yang berasal dari keton yang tidak simetris bersifat prokiral , reduksi mereka adalah metode yang berguna untuk sintesis amina kiral.
Sebagai ligan
Imina adalah ligan umum dalam kimia koordinasi . Kondensasi salicylaldehyde dan ethylenediamine memberi keluarga agen chelating yang mengandung imine seperti salen .
Pengurangan imina
Imina dapat direduksi menjadi amina melalui hidrogenasi misalnya dalam sintesis m -tolylbenzylamine: [16]
Asymmetric imine reduction pertama dilaporkan pada tahun 1973 oleh Kagan menggunakan Ph (Me) C = NBn dan PhSiH 2 dalam hidrosililasi dengan ligan kiral Katalis DIOP dan rhodium (RhCl (CH 2 CH 2 ) 2 ) 2 . [18] Banyak sistem telah diselidiki. [19] [20]
Biasanya imina mengacu pada senyawa dengan konektivitas R 2 C = NR, seperti yang dibahas di bawah ini. Dalam literatur yang lebih tua, imina mengacu pada analog aza dari epoksida. Dengan demikian, etilenimin adalah spesies cincin beranggota tiga C2H4 NH. [6]
Imina terkait dengan keton dan aldehid dengan penggantian oksigen dengan kelompok NR. Ketika R = H, senyawa tersebut adalah imina primer, ketika R adalah hidrokarbil , senyawa tersebut adalah imina sekunder. Imina menunjukkan reaktivitas yang beragam dan biasanya ditemui di seluruh kimia. [4] Ketika R 3 adalah OH, imine disebut oksime , dan ketika R 3 adalah NH 2 imina disebut hidrazon .
Imina primer di mana C melekat pada hidrokarbil dan H disebut aldimine primer ; imina sekunder dengan kelompok seperti ini disebut aldimine sekunder . [7] Imina primer di mana C melekat pada dua hidrokarbil disebut ketimine primer ; imina sekunder dengan kelompok seperti ini disebut ketimin sekunder . [8]
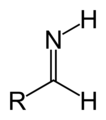 Aldimine primer
Aldimine primer
-skeletal.png&xid=17259,15700023,15700124,15700149,15700186,15700191,15700201,15700214&usg=ALkJrhgVaC_0vBVHGNKVzFwga-LDpwYdDw)
 Aldimine sekunder
Aldimine sekunder
-skeletal.png&xid=17259,15700023,15700124,15700149,15700186,15700191,15700201,15700214&usg=ALkJrhiic-SCkMmTr5Rqflj_8zTHrzTkRA)
 Ketimin primer
Ketimin primer
-skeletal.png&xid=17259,15700023,15700124,15700149,15700186,15700191,15700201,15700214&usg=ALkJrhhed_Q36gloRb494EMR2ggKyIgJbw)
 Ketimin sekunder
Ketimin sekunder
-skeletal.png&xid=17259,15700023,15700124,15700149,15700186,15700191,15700201,15700214&usg=ALkJrhiourqAF3W_ZjorUHIx-qAKMvCKPA)
 Aziridine dan turunannya kadang-kadang disebut sebagai imina.
Aziridine dan turunannya kadang-kadang disebut sebagai imina.

N-Sulfinyl imines adalah kelas khusus imina yang memiliki gugus sulfinil yang terikat pada atom nitrogen.
Sintesis imina
Imina biasanya disiapkan oleh kondensasi amina primer dan aldehida dan keton kurang umum:
- RNH 2 + R′C (O) R ′ ′ → RN = C (R ′) (R ′ ′) + H 2 O
Lebih banyak metode khusus
Beberapa metode lain ada untuk sintesis imina.
- Reaksi dari azida organik dengan logam carbenoid (diproduksi dari senyawa diazocarbonyl). [12]
- Kondensasi asam karbon dengan senyawa nitroso .
- Penataan ulang trityl N-haloamines dalam penataan ulang Stieglitz .
- Dehidrasi hemiaminal . [13]
- Dengan reaksi alkena dengan asam hidrazoat dalam reaksi Schmidt .
- Dengan reaksi asam nitril, asam klorida, dan aren dalam reaksi Hoesch .
- Sintesis multikomponen 3- tiazolina dalam reaksi Asinger .
- Ketimin primer dapat disintesis melalui reaksi Grignard dengan nitril . [14] [15]

Hexafluoroacetone imine adalah ketimine primer yang tidak biasa yang stabil.
- Imina berkurang dalam aminasi reduktif .
- Imina bereaksi dengan amina ke aminal , lihat misalnya sintesis cucurbituril .
- Sebuah imina bereaksi dengan diena dalam reaksi Aza Diels-Alder ke tetrahidropiridin.
- Imina dapat dioksidasi dengan asam meta-kloroperoksibenzoat (mCPBA) untuk memberikan oksaziridin
- Sebuah imina aromatik bereaksi dengan enol eter ke quinoline dalam reaksi Povarov .
- A tosylimine bereaksi dengan senyawa karbonil α, β-unsaturated ke amina alilik pada reaksi Aza-Baylis-Hillman .
- Imina adalah perantara dalam alkilasi amina dengan asam format dalam reaksi Eschweiler-Clarke .
- Penataan ulang dalam kimia karbohidrat yang melibatkan imina adalah penataan ulang Amadori .
- Reaksi transfer metilena dari imina oleh sulfonium ylide tidak stabil dapat memberikan sistem aziridine .
- Imina bereaksi, secara termal, dengan ketena dalam sikdisi siklik [2 + 2] untuk membentuk β-laktam dalam sintesis Staudinger .
- Sebuah imina dapat bereaksi dengan dialkilfosfit dalam reaksi Pudovik dan reaksi Kabachnik-Fields
Agak seperti induk amina, imina sedikit bersifat basa dan dapat berefleksi secara reversibel untuk menghasilkan garam iminium. Turunan iminium sangat rentan terhadap reduksi ke amina menggunakan transfer hidrogenasi atau dengan aksi stoikiometrik natrium cyanoborohidrida . Karena imina yang berasal dari keton yang tidak simetris bersifat prokiral , reduksi mereka adalah metode yang berguna untuk sintesis amina kiral.
Sebagai ligan
Imina adalah ligan umum dalam kimia koordinasi . Kondensasi salicylaldehyde dan ethylenediamine memberi keluarga agen chelating yang mengandung imine seperti salen .
Pengurangan imina
Imina dapat direduksi menjadi amina melalui hidrogenasi misalnya dalam sintesis m -tolylbenzylamine: [16]

Asymmetric imine reduction pertama dilaporkan pada tahun 1973 oleh Kagan menggunakan Ph (Me) C = NBn dan PhSiH 2 dalam hidrosililasi dengan ligan kiral Katalis DIOP dan rhodium (RhCl (CH 2 CH 2 ) 2 ) 2 . [18] Banyak sistem telah diselidiki. [19] [20]
Modifikasi
kimia dari protein tiol dengan reduksi dan alkilasi adalah umum dalam
persiapan sampel proteomik untuk analisis dengan spektrometri massa
(MS). Modifikasi pada kelompok fungsional lainnya kurang mendapat perhatian pada proteomik berbasis-MS.
Modifikasi amina (Lys, N-termini) oleh dimetilasi reduktif atau dengan
asilasi (misalnya pelabelan iTRAQ) baru-baru ini mendapatkan popularitas
dalam pendekatan berbasis peptida (MS dari bawah). Modifikasi pada kelompok asam (Asp, Glu, C-termini) telah dieksplorasi sangat minim.
Di sini, kami menggambarkan strategi pelabelan berurutan yang
memungkinkan modifikasi lengkap tiol, amina, dan asam pada peptida atau
protein utuh kecil. Metode ini termasuk ( 1 ) reduksi dan alkilasi tiol, ( 2 ) reduksi dimetilasi amina, dan ( 3 ) penengahan asam dengan beberapa amina.
Skema modifikasi kimia ini menawarkan beberapa pilihan baik untuk
penggabungan isotop stabil untuk kuantifikasi relatif dan untuk
meningkatkan peptida atau protein sebagai analit MS.
Modifikasi Grup Amino
 Modifikasi gugus amino melibatkan penambahan gugus fungsi pada ujung N dari asam amino .
Modifikasi gugus amino melibatkan penambahan gugus fungsi pada ujung N dari asam amino .
Protein setelah terjemahannya mengalami modifikasi kimia yang disebut Post Translational Modification . Modifikasi ini dapat mengubah fungsi protein ketika melekat pada kelompok fungsional biokimia seperti asetat dengan mengubah sifat kimia dari asam amino atau dengan perubahan struktural seperti lipat, distribusi konformasi, stabilitas, aktivitas dll.
Jenis Modifikasi Grup Amino
Asetilasi
Asetalisasi adalah proses asilasi (pengantar gugus asil ke senyawa organik) yang melibatkan substitusi gugus asam asetat organik untuk atom hidrogen aktif pada N-terminus.
Modifikasi paling luas pada eukariota adalah Asetilasi dari kelompok protein α-amina N-terminal. Sekitar 50% protein ragi dan sekitar 90% protein pada manusia dimodifikasi oleh mekanisme ini. Pola modifikasi dilestarikan sepanjang evolusi. Meskipun itu adalah modifikasi umum, tidak banyak informasi yang tersedia untuk tentang fungsi biologis dari asetilasi N-α-terminal. N-α-acetyltransferase (NATs) adalah enzim yang bertanggung jawab untuk Asetilasi. NAT milik keluarga GNAT, yang terletak di bawah superfamili acetyltransferases . [1]
 Asetilasi dan deasetilasi terjadi pada residu lisin di ekor N-terminal di asetilasi dan deasetilasi histone . Reaksi-reaksi ini terjadi di hadapan enzim histone acetyltransferase (HAT) atau histone deacetylase (HDAC).
Asetilasi dan deasetilasi terjadi pada residu lisin di ekor N-terminal di asetilasi dan deasetilasi histone . Reaksi-reaksi ini terjadi di hadapan enzim histone acetyltransferase (HAT) atau histone deacetylase (HDAC).
Formasi Pyroglutamate
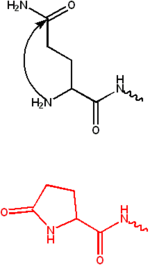 Pyroglutamate terbentuk melalui siklisasi (pembentukan cincin dalam senyawa kimia) glutamin . Ini biasanya diamati pada antibodi yang mengandung residu glutamat atau glutamin pada N-termini. Gugus amino dan glutamat atau glutamin mengembun membentuk cincin lima anggota yang disebut Pyroglutamate. Residu ini membuat protein lebih tahan terhadap aminopeptidase dan memiliki banyak peran fungsional. [2]
Pyroglutamate terbentuk melalui siklisasi (pembentukan cincin dalam senyawa kimia) glutamin . Ini biasanya diamati pada antibodi yang mengandung residu glutamat atau glutamin pada N-termini. Gugus amino dan glutamat atau glutamin mengembun membentuk cincin lima anggota yang disebut Pyroglutamate. Residu ini membuat protein lebih tahan terhadap aminopeptidase dan memiliki banyak peran fungsional. [2]
Ia mempertahankan integritas struktural pada N-terminal α-helix dan menyediakan lingkungan yang tepat untuk ionisasi residu Histidin untuk katalisis dan sitotoksisitas terhadap sel HeLa .
Myristoylation
N-myristoylation juga merupakan proses asilasi yang ditemukan khusus untuk gliserin asam amino N-terminal dalam protein di mana kelompok myristoy (berasal dari asam miristat) secara kovalen dilampirkan melalui ikatan amida dengan gugus alpha-amino N-terminal Glycine. Myristoylation memainkan peran penting dalam pensinyalan seluler sekunder, dalam infektivitas retrovirus dan onkogenesis pada Eukariota. Ini juga mempengaruhi fungsi fisiologis protein pengikat kalsium. [3] Enzim cytosolic N-myristoyltransferase (NMT) mengkatalisis Myristoylation.
Methylation
Metilasi protein adalah bentuk modifikasi pasca-translasi yang paling umum yang diamati. Mirip dengan modifikasi pasca-translasi lainnya, metilasi protein terlibat dalam mengatur interaksi protein-protein yang menghasilkan sejumlah besar efek selama peristiwa seluler utama, termasuk pengaturan transkripsi [4] [5] [6] respon stres, penuaan dan perbaikan protein [ 7] Aktivasi sel-T [8] , transportasi nuklir [9] , diferensiasi neuronal [10] , [11] fungsi saluran ion, dan pensinyalan sitokin. Protein dianggap termetilasi ketika kelompok metil ditambahkan pada satu atau lebih rantai samping nukleofilik. Metilasi pada rantai samping nitrogens dianggap sangat tidak dapat diubah sementara metilasi dari gugus karboksil berpotensi reversibel. Residu protein yang termetilasi pada nitrogen termasuk e-amin dari lisin, cincin imidazol histidin, bagian guanidino arginin, dan rantai samping amida nitrit glutamin dan asparagin.
 Metilasi dalam protein meniadakan muatan negatif di atasnya dan meningkatkan hidrofobisitas protein. Metilasi pada rantai samping karboksilat menutupi muatan negatif dan menambah hidrofobik. N-Metilasi lisin tidak mengubah muatan kationik tetapi meningkatkan hidrofobik. Khususnya, dimetilasi dan trimetilasi rantai samping lisin
dalam protein meningkatkan baik hidrofobik dan massa sterik dan dapat
mempengaruhi interaksi protein-protein jika mereka berada dalam
permukaan yang berinteraksi.
Metilasi dalam protein meniadakan muatan negatif di atasnya dan meningkatkan hidrofobisitas protein. Metilasi pada rantai samping karboksilat menutupi muatan negatif dan menambah hidrofobik. N-Metilasi lisin tidak mengubah muatan kationik tetapi meningkatkan hidrofobik. Khususnya, dimetilasi dan trimetilasi rantai samping lisin
dalam protein meningkatkan baik hidrofobik dan massa sterik dan dapat
mempengaruhi interaksi protein-protein jika mereka berada dalam
permukaan yang berinteraksi.
Karbamilasi
Karbamilasi terjadi ketika asam isocyanic (HCNO) bereaksi dengan residu amino terminus, seperti lisin, dari protein. Ini adalah salah satu modifikasi protein artifactual umum yang diakui untuk fokus Isoeletric . Faktor risikonya adalah urea (chaotrope) yang ada dalam larutan dan berada dalam kesetimbangan dengan amonium sianat . Asam isosianat adalah bentuk sianat yang bereaksi dengan gugus amino protein. Untuk karbamilasi terjadi gugus asam amino protein seperti lisin, rantai samping arginin harus terdeprotonasi yang biasanya terjadi pada pH basa. Karbamilasi terjadi ketika protein dibiarkan pada suhu kamar dalam larutan urea dan di mana asam isosianat dapat dengan bebas bereaksi dengan protein.

Karbamilasi oleh asam isosianat negatif untuk langkah selanjutnya dari karakterisasi protein karena asam isosianat bereaksi dengan ujung amino protein memblokir peptida atau protein ke sekuens N-terminal. Asam isosianat menyerang rantai samping residu lisin dan arginin yang membuat protein tidak cocok untuk banyak pencernaan enzim. Bahkan jika karbamilasi tidak mencegah digest enzimatik, seringkali akan membingungkan hasil dari eksperimen spektroskopi massa dengan peptida yang memiliki waktu retensi dan massa yang tidak terduga. Karbamilasi protein in vivo diamati di beberapa negara berpenyakit. [12]
Formilasi
Formilasi adalah salah satu modifikasi posttranslational protein, di mana protein dimodifikasi oleh lampiran kelompok formil. Mekanisme yang paling umum dipelajari adalah N6-formilasi lisin yang terkait dengan histone dan protein nuklir lainnya. Modifikasi translasi post histone dan protein kromatin lainnya memainkan peran dalam fisiologi ekspresi gen . "Residu N6-formil-lisin muncul untuk mewakili modifikasi sekunder histone endogen, yang mengandung kesamaan kimia dengan lisin N6-asetilasi yang diakui sebagai penentu penting ekspresi gen pada sel mamalia." Dari penelitian itu disimpulkan bahwa modifikasi N6-formil dari lisin mengganggu fungsi sinyal asetilasi dan metilasi, yang memainkan peran dalam fisiologi stres oksidatif dan nitrosatif. [13]

Protein setelah terjemahannya mengalami modifikasi kimia yang disebut Post Translational Modification . Modifikasi ini dapat mengubah fungsi protein ketika melekat pada kelompok fungsional biokimia seperti asetat dengan mengubah sifat kimia dari asam amino atau dengan perubahan struktural seperti lipat, distribusi konformasi, stabilitas, aktivitas dll.
Jenis Modifikasi Grup Amino
Asetilasi
Asetalisasi adalah proses asilasi (pengantar gugus asil ke senyawa organik) yang melibatkan substitusi gugus asam asetat organik untuk atom hidrogen aktif pada N-terminus.
Modifikasi paling luas pada eukariota adalah Asetilasi dari kelompok protein α-amina N-terminal. Sekitar 50% protein ragi dan sekitar 90% protein pada manusia dimodifikasi oleh mekanisme ini. Pola modifikasi dilestarikan sepanjang evolusi. Meskipun itu adalah modifikasi umum, tidak banyak informasi yang tersedia untuk tentang fungsi biologis dari asetilasi N-α-terminal. N-α-acetyltransferase (NATs) adalah enzim yang bertanggung jawab untuk Asetilasi. NAT milik keluarga GNAT, yang terletak di bawah superfamili acetyltransferases . [1]

Formasi Pyroglutamate

Ia mempertahankan integritas struktural pada N-terminal α-helix dan menyediakan lingkungan yang tepat untuk ionisasi residu Histidin untuk katalisis dan sitotoksisitas terhadap sel HeLa .
Myristoylation
N-myristoylation juga merupakan proses asilasi yang ditemukan khusus untuk gliserin asam amino N-terminal dalam protein di mana kelompok myristoy (berasal dari asam miristat) secara kovalen dilampirkan melalui ikatan amida dengan gugus alpha-amino N-terminal Glycine. Myristoylation memainkan peran penting dalam pensinyalan seluler sekunder, dalam infektivitas retrovirus dan onkogenesis pada Eukariota. Ini juga mempengaruhi fungsi fisiologis protein pengikat kalsium. [3] Enzim cytosolic N-myristoyltransferase (NMT) mengkatalisis Myristoylation.
Methylation
Metilasi protein adalah bentuk modifikasi pasca-translasi yang paling umum yang diamati. Mirip dengan modifikasi pasca-translasi lainnya, metilasi protein terlibat dalam mengatur interaksi protein-protein yang menghasilkan sejumlah besar efek selama peristiwa seluler utama, termasuk pengaturan transkripsi [4] [5] [6] respon stres, penuaan dan perbaikan protein [ 7] Aktivasi sel-T [8] , transportasi nuklir [9] , diferensiasi neuronal [10] , [11] fungsi saluran ion, dan pensinyalan sitokin. Protein dianggap termetilasi ketika kelompok metil ditambahkan pada satu atau lebih rantai samping nukleofilik. Metilasi pada rantai samping nitrogens dianggap sangat tidak dapat diubah sementara metilasi dari gugus karboksil berpotensi reversibel. Residu protein yang termetilasi pada nitrogen termasuk e-amin dari lisin, cincin imidazol histidin, bagian guanidino arginin, dan rantai samping amida nitrit glutamin dan asparagin.

Karbamilasi
Karbamilasi terjadi ketika asam isocyanic (HCNO) bereaksi dengan residu amino terminus, seperti lisin, dari protein. Ini adalah salah satu modifikasi protein artifactual umum yang diakui untuk fokus Isoeletric . Faktor risikonya adalah urea (chaotrope) yang ada dalam larutan dan berada dalam kesetimbangan dengan amonium sianat . Asam isosianat adalah bentuk sianat yang bereaksi dengan gugus amino protein. Untuk karbamilasi terjadi gugus asam amino protein seperti lisin, rantai samping arginin harus terdeprotonasi yang biasanya terjadi pada pH basa. Karbamilasi terjadi ketika protein dibiarkan pada suhu kamar dalam larutan urea dan di mana asam isosianat dapat dengan bebas bereaksi dengan protein.

Karbamilasi oleh asam isosianat negatif untuk langkah selanjutnya dari karakterisasi protein karena asam isosianat bereaksi dengan ujung amino protein memblokir peptida atau protein ke sekuens N-terminal. Asam isosianat menyerang rantai samping residu lisin dan arginin yang membuat protein tidak cocok untuk banyak pencernaan enzim. Bahkan jika karbamilasi tidak mencegah digest enzimatik, seringkali akan membingungkan hasil dari eksperimen spektroskopi massa dengan peptida yang memiliki waktu retensi dan massa yang tidak terduga. Karbamilasi protein in vivo diamati di beberapa negara berpenyakit. [12]
Formilasi
Formilasi adalah salah satu modifikasi posttranslational protein, di mana protein dimodifikasi oleh lampiran kelompok formil. Mekanisme yang paling umum dipelajari adalah N6-formilasi lisin yang terkait dengan histone dan protein nuklir lainnya. Modifikasi translasi post histone dan protein kromatin lainnya memainkan peran dalam fisiologi ekspresi gen . "Residu N6-formil-lisin muncul untuk mewakili modifikasi sekunder histone endogen, yang mengandung kesamaan kimia dengan lisin N6-asetilasi yang diakui sebagai penentu penting ekspresi gen pada sel mamalia." Dari penelitian itu disimpulkan bahwa modifikasi N6-formil dari lisin mengganggu fungsi sinyal asetilasi dan metilasi, yang memainkan peran dalam fisiologi stres oksidatif dan nitrosatif. [13]
Amina adalah senyawa organik
dan gugus fungsi yang mengandung nitrogen basa dengan pasangan elektron
bebas. Perlindungan nitrogen terus menarik banyak perhatian dalam
bidang kimia, seperti peptida, nukleosida, polimer dan sintesis ligan.
Tetapi, dalam beberapa tahun terakhir, sejumlah gugus pelindung nitrogen
telah digunakan sebagai pembantu kiral. Dengan demikian, desain baru,
lebih ringan dan metodenya lebih efektif untuk perlindungan nitrogen
masih aktif dalam topik sintesis kimia.

Gugus
Pelindung imida dan amida: Kelompok ftalimida telah berhasil digunakan
untuk melindungi gugus amino. Pembelahan dari N-alkilftalimida (1,81)
mudah dilakukan dengan hidrazin, dalam larutan panas atau dalam dingin
untuk waktu yang lama untuk memberikan (1,82) dan amina. Basa katalis
hidrolisis N-alkilftalimida (1.81) juga memberikan yang sesuai amina.

Modifikasi Kitosan
Adanya gugus amina (NH2) dan dan hidroksil (OH) dari kitosan menyebabkan kitosan mudah dimodifikasi secara kimia.

Gugus aktif pada kitosan
Bila
dibutuhkan perubahan gugus fungsional untuk menghalangi gangguan dalam
beberapa rangkaian reaksi sintesis, salah satu caranya adalah dengan
menggunakan gugus pelindung. Gugus pelindung merupakan suatu turunan
yang dapat dibuat dan kemudian dihilangkan. Tiga syarat-syarat yang
perlu diperhatikan dalam memilih gugus pelindung adalah sebagai berikut :
1. Gugus pelindung yang digunakan harus lebih reaktif
2. Gugus pelindung yang dipakai harus dengan mudah bereaksi dengan molekul target.
3. Kondisi reaksi dalam memasukkan gugus pelindung harus stabil.
4. Dapat dimasukkan pada kondisi reaksi lunak
5. Gugus pelindung harus dapat dengan mudah dihilangkan tanpa menggangu reaksi akhir.
Reaksi
penggunaan gugus pelindung pada kitosan dikarenakan kitosan memiliki 2
gugus fungsi yang kereaktifan berbeda. Gugus amino dari kitosan lebih
reaktif dari pada gugus hidroksilnya, sehingga untuk menghasilkan
O-asilasi kitosan, perlu dilakukan proteksi atau perlindungan terhadap
gugus amino. Basa shiff dapat digunakan sebagai gugus pelindung pada
reaksi O-asilasi.
Gugus
amino kitosan lebih reaktif daripada gugus hidroksilnya, sehingga untuk
menghasilkan O-asilasi kitosan perlu dilakukan proteksi atau
perlindungan terhadap gugus amin selama proses asilasi untuk
menghasilkan O-Asil kitosan. Metode proteksi yang dilakukan antara lain
melalui pembuatan basa Schiff disusul O-Asetilasi menggunakan larutan
asetat anhidrin-piridin untuk mencegah hidrolisis asam dari basa Schiff.
Reaksi antara kitosan dengan anhidrida asetat menghasilkan senyawa
ester yang merupakan kitosan asetat. Dalam hal ini kitosan terlebih
dahulu direaksikan dengan asetaldehida membentuk aldimin untuk
melindungi gugus amina. Kitosan laurat diperoleh dari reaksi
transesterifikasi antara metil laurat dengan kitosan asetat. Selanjutnya
dilakukan deproteksi dengan menambahkan natrium bikarbonat untuk
memperoleh kitosan laurat.


Basa Schiff dapat digunakan sebagai gugus pelindung pada gugus amin (NH2),
dilakukan dengan melarutkan kitosan terasetilasi dalam asam formiat 90%
yang mengandung asetat anhidrida dengan asumsi protonasi akan mencegah
terjadinya N-asilasi. Selanjutnya direaksikan dengan asilklorida dalam
karbon triklorida dan piridin kering.
Contoh lain gugus pelindung untuk NH2 yaitu:

peramasalahan
1. Bagaimana sintesis imin dengan penataan ulang trityl N-haloamines dalam penataan ulang Stieglitz ?
2. Bagaimana Modifikasi translasi post histone dan protein kromatin lainnya memainkan peran dalam fisiologi ekspresi gen ?
3. Protein setelah terjemahannya mengalai modifikasi kimia yang disebut Post Translational Modification . Modifikasi ini dapat mengubah fungsi protein ketika melekat pada kelompok fungsional biokimia seperti asetat dengan mengubah sifat kimia dari asam amino atau dengan perubahan struktural seperti lipat, distribusi konformasi, stabilitas, aktivitas dll.Bagaimana proses terjadinya didalam tubuh ?


No 2 Epigenetik adalah mekanisme intrinsik yang merubah ekspresi gen bukan dengan merubah sekuens DNA tetapi dengan memodifikasi DNA dan protein kromosom histon. Epigenetik terlibat dalam genomic imprinting dan aktivasi kromosom X pada manusia dan kegagalan dari mekanisme ini menyebabkan kelainan kongenital dan timbulnya kanker (Kubota, 2008).
ReplyDeleteTerjadinya proses epigenetik (epigenesis) merupakan konsekuensi adanya interaksi antara gen dan lingkungannya dan dapat terjadi akibat tidak terekspresinya informasi genomik (silenced genes). Epigenesis mencakup seluruh mekanisme yang menyebabkan perbedaan ekspresi gen pada sel-sel tertentu. Pengaturan epigenetik ditentukan oleh : metilasi DNA, modifikasi histon, ekspresi gen miRNA, faktor transkripsi jaringan (Egger, 2004).Mekanisme tersebut mencakup metilasi DNA, konfigurasi kromatin, atau kombinasi keduanya. Konsekuensi dari mekanisme tersebut adalah sebuah spektrum gen-gen yang aktif dan gen-gen yang tidak aktif (silent) pada setiap tipe sel yang ada. Epigenesis juga meliputi mekanisme yang bertanggung jawab menentukan program genetik untuk perkembangan (development) di mana mekanisme ini bergantung pada proses-proses seperti pensinyalan sel dan berbagai interaksi seluler lainnya. Epigenesis juga bertanggung jawab atas terjadinya genomic imprinting di mana beberapa gen yang berasal dari gamet jantan dan betina memiliki ekspresi yang berbeda
Struktur kromatin ditentukan oleh DNA metilasi dan modifikasi pada protein histon. Mekanisme ini merupakan mekanisme utama epigenetik untuk mengontrol ekspresi gen.Metilasi DNA adalah salah satu modifikasi epigenetik yang tidak merubah urutan sekuense utama DNA, namun merupakan faktor kritis bagi perkembangan yang normal, pola ekspresi gen, dan stabilitas genomik, dengan cara penambahan kovalen gugus metil pada ujung 5’ sitosin dalam CpG dinukleotida dan CpNpG trinukleotida yang sebagian besar berkelompok pada daerah genom yang disebut sebagai CpG island. Proses ini difasilitasioleh suatu enzim yaitu DNA metiltransferase (DNMTs). Secara umum, hipo dan hipermetilasi berhubungan dengan ekspresi gen dan silent gen. Pola metilasi DNA diprakarsai dan dikelola oleh DNMTs
Struktur kromatin ditentukan oleh DNA metilasi dan modifikasi pada protein histon. Mekanisme ini merupakan mekanisme utama epigenetik untuk mengontrol ekspresi gen.Metilasi DNA adalah salah satu modifikasi epigenetik yang tidak merubah urutan sekuense utama DNA, namun merupakan faktor kritis bagi perkembangan yang normal, pola ekspresi gen, dan stabilitas genomik, dengan cara penambahan kovalen gugus metil pada ujung 5’ sitosin dalam CpG dinukleotida dan CpNpG trinukleotida yang sebagian besar berkelompok pada daerah genom yang disebut sebagai CpG island. Proses ini difasilitasioleh suatu enzim yaitu DNA metiltransferase (DNMTs). Secara umum, hipo dan hipermetilasi berhubungan dengan ekspresi gen dan silent gen. Pola metilasi DNA diprakarsai dan dikelola oleh DNMTs
ReplyDeleteSaya akan mencoba menjawab permasalahan no 1
ReplyDeleteSalah satu cara menamai aldimines adalah dengan mengambil nama radikal, hapus "e" akhir, dan tambahkan "-imine", misalnya ethanimine . Lihat artikel aldimine untuk konvensi penamaan lainnya. Secara bergantian, sebuah imina dinamakan sebagai turunan dari karbonil, menambahkan kata "imina" ke nama senyawa karbonil yang gugus oksonya digantikan oleh kelompok imino , misalnya imon sydnone dan imina aseton (zat antara dalam sintesis). acetone azine ).
N-Sulfinyl imines adalah kelas khusus imina yang memiliki gugus sulfinil yang terikat pada atom nitrogen.
Sintesis imina
Imina biasanya disiapkan oleh kondensasi amina primer dan aldehida dan keton kurang umum:
RNH 2 + R′C (O) R ′ ′ → RN = C (R ′) (R ′ ′) + H 2 O
Dalam hal mekanisme, reaksi tersebut dilanjutkan melalui penambahan nukleofilik memberikan hemiaminal -C (OH) (NHR) - menengah, diikuti dengan eliminasi air untuk menghasilkan imina. (lihat alkylimino-de-oxo-bisubstitution untuk mekanisme terperinci) Kesetimbangan dalam reaksi ini biasanya mendukung senyawa karbonil dan amina, sehingga distilasi azeotropik atau penggunaan agen dehidrasi, seperti saringan molekuler atau magnesium sulfat , diperlukan untuk mendorong reaksi yang mendukung pembentukan imina. Dalam beberapa tahun terakhir, beberapa reagen seperti Tris (2,2,2-trifluoroethyl) borat [B (OCH 2 CF 3 ) 3 ], [9] pyrrolidine [10] atau titanium ethoxide [Ti (OEt) 4 ] [11] telah terbukti mengkatalisis pembentukan imina.
Saya akan menjawab no 3.
ReplyDeleteModifikasi translasi pasca terjadi pada ujung peptida dari rantai asam amino memainkan peran penting dalam mentranslokasi mereka di seluruh membran biologis. Ini termasuk protein sekretorik dalam prokariota dan eukariota dan juga protein yang dimaksudkan untuk dimasukkan dalam berbagai membran seluler dan organel seperti lisosom, kloroplas, mitokondria dan membran plasma.
Ekspresi protein penting dalam kondisi berpenyakit. Modifikasi translasi pasca memainkan bagian penting dalam memodifikasi produk akhir dari ekspresi dan berkontribusi terhadap proses biologis dan kondisi yang sakit. Urutan terminal amino dihapus oleh pembelahan proteolitik ketika protein melintasi membran. Sekuens-sekuen terminal amino ini mentargetkan protein untuk mengangkutnya ke titik aksi yang sebenarnya di dalam sel.
Protein pasca translasi modifikasi dapat terjadi dalam beberapa cara. Beberapa dari mereka terdaftar di bawah ini:
Glikosilasi: Banyak protein, terutama pada sel eukariotik, dimodifikasi oleh penambahan karbohidrat, suatu proses yang disebut glikosilasi. Glikosilasi pada protein menghasilkan tambahan gugus glikosil baik pada asparagin, hidroksisin, serin, atau treonin. Perangkat lunak untuk mempelajari glikosilasi oleh prediksi struktur glycan.
Asetilasi: penambahan gugus asetil, biasanya pada N-terminus protein.
Alkilasi: Penambahan gugus alkil (misalnya metil, etil).
Metilasi: Penambahan kelompok metil, biasanya pada residu lisin atau arginin. (Ini adalah jenis alkilasi.)
Biotinylation: Asilasi residu lisin yang dilestarikan dengan tambahan biotin.
Glutamilasi: Hubungan konvertal residu asam glutamat ke tubulin dan beberapa protein lainnya.
Glycylation: Hubungan kovalen dari satu hingga lebih dari 40 residu glisin ke tubulin C-terminal ekor dari rangkaian asam amino.
Isoprenylation: Penambahan kelompok isoprenoid (misalnya farnesol dan geranylgeraniol).
Lipoylation: Keterikatan fungsi lipoat.
Phosphopantetheinylation, Penambahan bagian 4'-phosphopantetheinyl dari koenzim A, seperti pada asam lemak, poliketida, peptida non-ribosom dan biosintesis leusin.
Fosforilasi, penambahan gugus fosfat, biasanya menjadi serin, tyrosin, treonin atau histidin.
Sulfasi: Penambahan kelompok sulfat ke tirosin.
Selenasi
Penengahan C-terminal
baiklah saya akan mencoba menjawab permasalahan nomor 2
ReplyDeleteNo 2 dimana yaitu Epigenetik adalah mekanisme intrinsik yang merubah ekspresi gen bukan dengan merubah sekuens DNA tetapi dengan memodifikasi DNA dan protein kromosom histon. Epigenetik terlibat dalam genomic imprinting dan aktivasi kromosom X pada manusia dan kegagalan dari mekanisme ini menyebabkan kelainan kongenital dan timbulnya kanker (Kubota, 2008).
Terjadinya proses epigenetik (epigenesis) merupakan konsekuensi adanya interaksi antara gen dan lingkungannya dan dapat terjadi akibat tidak terekspresinya informasi genomik (silenced genes). Epigenesis mencakup seluruh mekanisme yang menyebabkan perbedaan ekspresi gen pada sel-sel tertentu. Pengaturan epigenetik ditentukan oleh : metilasi DNA, modifikasi histon, ekspresi gen miRNA, faktor transkripsi jaringan (Egger, 2004).Mekanisme tersebut mencakup metilasi DNA, konfigurasi kromatin, atau kombinasi keduanya. Konsekuensi dari mekanisme tersebut adalah sebuah spektrum gen-gen yang aktif dan gen-gen yang tidak aktif (silent) pada setiap tipe sel yang ada. Epigenesis juga meliputi mekanisme yang bertanggung jawab menentukan program genetik untuk perkembangan (development) di mana mekanisme ini bergantung pada proses-proses seperti pensinyalan sel dan berbagai interaksi seluler lainnya. Epigenesis juga bertanggung jawab atas terjadinya genomic imprinting di mana beberapa gen yang berasal dari gamet jantan dan betina memiliki ekspresi yang berbeda.
Saya akan mencoba menjawab permasalahan yang ke 2Imina biasanya disiapkan oleh kondensasi amina primer dan aldehida dan keton kurang umum:
ReplyDeleteRNH 2 + R′C (O) R ′ ′ → RN = C (R ′) (R ′ ′) + H 2 O
Dalam hal mekanisme, reaksi tersebut dilanjutkan melalui penambahan nukleofilik memberikan hemiaminal -C (OH) (NHR) - menengah, diikuti dengan eliminasi air untuk menghasilkan imina. (lihat alkylimino-de-oxo-bisubstitution untuk mekanisme terperinci) Kesetimbangan dalam reaksi ini biasanya mendukung senyawa karbonil dan amina, sehingga distilasi azeotropik atau penggunaan agen dehidrasi, seperti saringan molekuler atau magnesium sulfat , diperlukan untuk mendorong reaksi yang mendukung pembentukan imina. Dalam beberapa tahun terakhir, beberapa reagen seperti Tris (2,2,2-trifluoroethyl) borat [B (OCH 2 CF 3 ) 3 ], [9] pyrrolidine [10] atau titanium ethoxide [Ti (OEt) 4 ] [11] telah terbukti mengkatalisis pembentukan imina.
Struktur kromatin ditentukan oleh DNA metilasi dan modifikasi pada protein histon. Mekanisme ini merupakan mekanisme utama epigenetik untuk mengontrol ekspresi gen.Metilasi DNA adalah salah satu modifikasi epigenetik yang tidak merubah urutan sekuense utama DNA, namun merupakan faktor kritis bagi perkembangan yang normal, pola ekspresi gen, dan stabilitas genomik, dengan cara penambahan kovalen gugus metil pada ujung 5’ sitosin dalam CpG dinukleotida dan CpNpG trinukleotida yang sebagian besar berkelompok pada daerah genom yang disebut sebagai CpG island. Proses ini difasilitasioleh suatu enzim yaitu DNA metiltransferase (DNMTs). Secara umum, hipo dan hipermetilasi berhubungan dengan ekspresi gen dan silent gen. Pola metilasi DNA diprakarsai dan dikelola oleh DNMTs
ReplyDeleteSaya akan menjawab no 3.
ReplyDeleteModifikasi translasi pasca terjadi pada ujung peptida dari rantai asam amino memainkan peran penting dalam mentranslokasi mereka di seluruh membran biologis. Ini termasuk protein sekretorik dalam prokariota dan eukariota dan juga protein yang dimaksudkan untuk dimasukkan dalam berbagai membran seluler dan organel seperti lisosom, kloroplas, mitokondria dan membran plasma.
Ekspresi protein penting dalam kondisi berpenyakit. Modifikasi translasi pasca memainkan bagian penting dalam memodifikasi produk akhir dari ekspresi dan berkontribusi terhadap proses biologis dan kondisi yang sakit. Urutan terminal amino dihapus oleh pembelahan proteolitik ketika protein melintasi membran. Sekuens-sekuen terminal amino ini mentargetkan protein untuk mengangkutnya ke titik aksi yang sebenarnya di dalam sel.
jawaban permasalahan no 1 : Basa Schiff dapat digunakan sebagai gugus pelindung pada gugus amin (NH2), dilakukan dengan melarutkan kitosan terasetilasi dalam asam formiat 90% yang mengandung asetat anhidrida dengan asumsi protonasi akan mencegah terjadinya N-asilasi. Selanjutnya direaksikan dengan asilklorida dalam karbon triklorida dan piridin kering.
ReplyDeleteNomor 2
ReplyDeleteStruktur kromatin ditentukan oleh DNA metilasi dan modifikasi pada protein histon. Mekanisme ini merupakan mekanisme utama epigenetik untuk mengontrol ekspresi gen.Metilasi DNA adalah salah satu modifikasi epigenetik yang tidak merubah urutan sekuense utama DNA, namun merupakan faktor kritis bagi perkembangan yang normal, pola ekspresi gen, dan stabilitas genomik, dengan cara penambahan kovalen gugus metil pada ujung 5’ sitosin dalam CpG dinukleotida dan CpNpG trinukleotida yang sebagian besar berkelompok pada daerah genom yang disebut sebagai CpG island. Proses ini difasilitasioleh suatu enzim yaitu DNA metiltransferase (DNMTs). Secara umum, hipo dan hipermetilasi berhubungan dengan ekspresi gen dan silent gen. Pola metilasi DNA diprakarsai dan dikelola oleh DNMTs
Saya akan mencoba menjawab permasalahan no 1
ReplyDeleteSalah satu cara menamai aldimines adalah dengan mengambil nama radikal, hapus "e" akhir, dan tambahkan "-imine", misalnya ethanimine . Lihat artikel aldimine untuk konvensi penamaan lainnya. Secara bergantian, sebuah imina dinamakan sebagai turunan dari karbonil, menambahkan kata "imina" ke nama senyawa karbonil yang gugus oksonya digantikan oleh kelompok imino , misalnya imon sydnone dan imina aseton (zat antara dalam sintesis). acetone azine ).
Nomor 2
ReplyDeleteStruktur kromatin ditentukan oleh DNA metilasi dan modifikasi pada protein histon. Mekanisme ini merupakan mekanisme utama epigenetik untuk mengontrol ekspresi gen.Metilasi DNA adalah salah satu modifikasi epigenetik yang tidak merubah urutan sekuense utama DNA, namun merupakan faktor kritis bagi perkembangan yang normal, pola ekspresi gen, dan stabilitas genomik, dengan cara penambahan kovalen gugus metil pada ujung 5’ sitosin dalam CpG dinukleotida dan CpNpG trinukleotida yang sebagian besar berkelompok pada daerah genom yang disebut sebagai CpG island. Proses ini difasilitasioleh suatu enzim yaitu DNA metiltransferase (DNMTs). Secara umum, hipo dan hipermetilasi berhubungan dengan ekspresi gen dan silent gen. Pola metilasi DNA diprakarsai dan dikelola oleh DNMTs